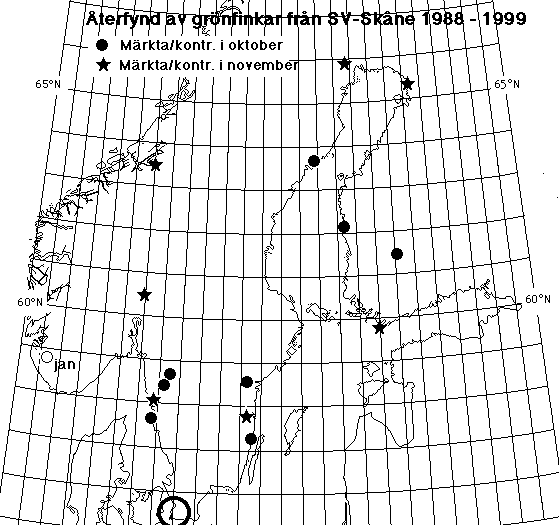
|
|
Ruggning och flyttning hos grönfinken (Carduelis chloris) i SV-Skåne |
| For readers from other continents: The Greenfinch Carduelis chloris is mainly a European species, breeding from North Africa to N Scandinavia. In addition it breeds in Asia from Turkey to Uzbekistan. It has a full postnuptial moult and winters in the breeding area at least to 56ş N in W Europe. |

| Period | n | Färdigruggade fåglar: n / % | Fåglar i pennruggning: n / % | Återstående medelscore hos ruggare ± 1 s.e.; 1 s.d. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 30.10 - 29.11.82 | 232 | 160 / 69.0 | 72 / 31.0 | 12.0 ± 1.4; 11.8 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 24..10 - 14.11.88 | 134 | 88 / 65.7 | 46 / 34.3 | 7.3 ± 1.2; 7.8 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 16.10 - 21.11.90 | 219 | 133 / 60.7 | 86 / 39.3 | 11.2 ± 1.3; 12.2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 14.10 - 16.11.94 | 210 | 99 / 47.1 | 111 / 52.9 | 10.6 ± 1.0; 10.5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 15.10 - 8.11.97 | 71 | 48 / 67.6 | 23 / 32.4 | 12.9 ± 2.0; 9.4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 16.10 - 3.11.99 | 130 | 95 / 73.1 | 35 / 26.9 | 10.1 ± 1.4; 8.4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
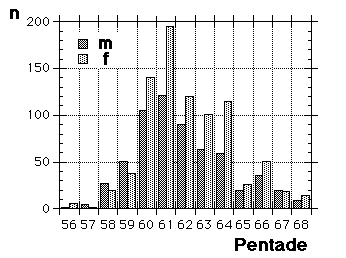
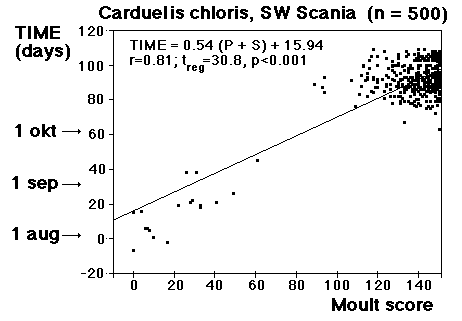
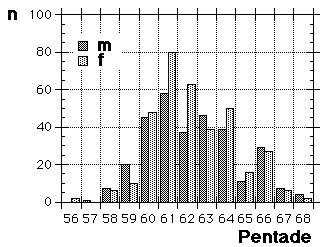
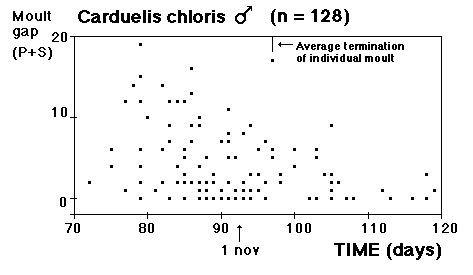
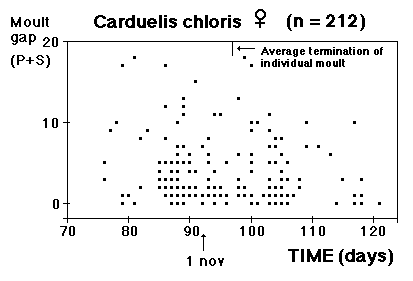
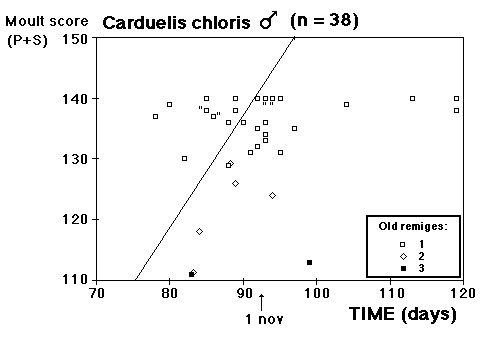
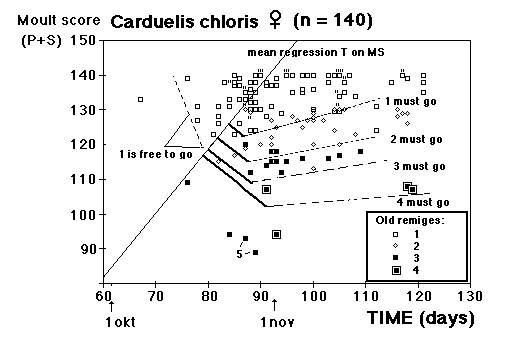
 Fig. 8 visar fördelningen för fåglar i minst sitt tredje kalenderår (3k+), fångade i Ljunghusen, Lund och Löddesnäs mellan 2.3 och 20.5 1979 - 2001. Medianen för hanarna infaller den 16 april, för honorna 13 april: observera att hanarna är i majoritet, men om man tar bort överskottet (30 hanar) i början av maj blir könsproportionerna 50 : 50 och medianen för bägge könen hamnar på den 13 april. I april uppträder de sträckande fåglarna ofta som distinkta par, hanen följer en speciell hona så fort hon flyger, det är vanligare med ensamma hanar i maj, vilket kunde tyda på att de flesta är lokala och har ruvande honor. (Härifrån borde man ändå gå ett steg vidare och kalkylera den förväntade livslängden för könen av olika åldersklasser hos grönfinken; materialet finns, man behöver inte ta till mer än de publicerade fynden).
Fig. 8 visar fördelningen för fåglar i minst sitt tredje kalenderår (3k+), fångade i Ljunghusen, Lund och Löddesnäs mellan 2.3 och 20.5 1979 - 2001. Medianen för hanarna infaller den 16 april, för honorna 13 april: observera att hanarna är i majoritet, men om man tar bort överskottet (30 hanar) i början av maj blir könsproportionerna 50 : 50 och medianen för bägge könen hamnar på den 13 april. I april uppträder de sträckande fåglarna ofta som distinkta par, hanen följer en speciell hona så fort hon flyger, det är vanligare med ensamma hanar i maj, vilket kunde tyda på att de flesta är lokala och har ruvande honor. (Härifrån borde man ändå gå ett steg vidare och kalkylera den förväntade livslängden för könen av olika åldersklasser hos grönfinken; materialet finns, man behöver inte ta till mer än de publicerade fynden).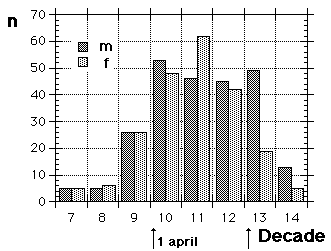 För att markera att jag går mot strömmen, att min tankeriktning är motsatt den darwinistiska riktningen, väljer jag att beteckna en del av honpopulationen hos grönfinken som lite "missanpassad" i sin ruggning, konsekvent, systematiskt missanpassad. En utmärkt term, utmanande i sin dubbla konnotation (den talar också om att fågeln inte kommer att dras genom det gängse adaptiva tröskverket, utan att jag siktar in mig på växelspelet mellan individerna och den dynamik som är ett resultat av detta växelspel). Detta är alltså min utgångspunkt: grönfinkhonans missanpassade ruggning.Avbrottet är alltså katastrofens viktigaste funktion. Men man får inte blunda för sekundära effekter: den kan också maka lätt på tidsgränser, i den mån det finns inre klockor, som löper från avbrottet av föregående års häckning eller ruggning (referenser här). Det är därför det kan vara fördelaktigt om en inre klocka mäter ut en period, som är något kortare än kalenderåret. På det viset blir katastrofen också kreativ, eller för att bruka en term från Levontin (1978): problemlösande (hos Levontin dock ej applicerat på katastroffunktionen). Framåtriktat kreativ, i en teleonom, vagt målsökande process.
För att markera att jag går mot strömmen, att min tankeriktning är motsatt den darwinistiska riktningen, väljer jag att beteckna en del av honpopulationen hos grönfinken som lite "missanpassad" i sin ruggning, konsekvent, systematiskt missanpassad. En utmärkt term, utmanande i sin dubbla konnotation (den talar också om att fågeln inte kommer att dras genom det gängse adaptiva tröskverket, utan att jag siktar in mig på växelspelet mellan individerna och den dynamik som är ett resultat av detta växelspel). Detta är alltså min utgångspunkt: grönfinkhonans missanpassade ruggning.Avbrottet är alltså katastrofens viktigaste funktion. Men man får inte blunda för sekundära effekter: den kan också maka lätt på tidsgränser, i den mån det finns inre klockor, som löper från avbrottet av föregående års häckning eller ruggning (referenser här). Det är därför det kan vara fördelaktigt om en inre klocka mäter ut en period, som är något kortare än kalenderåret. På det viset blir katastrofen också kreativ, eller för att bruka en term från Levontin (1978): problemlösande (hos Levontin dock ej applicerat på katastroffunktionen). Framåtriktat kreativ, i en teleonom, vagt målsökande process.