
|
|
Moult of the Whitethroat Sylvia communis in SW Scania |
| For readers from other continents: The (Common) Whitethroat Sylvia communis is a West Palearctic species, breeding to appr. 100ş E in Siberia. All populations winter in Africa and have a full postnuptial moult before or after the autumn migration. A few remiges may be exchanged before the spring migration, this applies to most Sylvia species. There are differences in suspension rate between W European populations and birds belonging to the eastern subspecies icterops. |
There is a tendency towards a faint "icterops moult pattern" in south Swedish Whitethroats; many birds will suspend some secondaries before autumn migration. Such suspension will shorten the duration of moult to little more than 30 days; it occurs in a two-brooded population near the northern limit for two broods. Rainfall and food abundance in the breeding areas in autumn as well as in Africa in winter, rather than migration distance, are thought to lie behind the exact shape of the moult strategy in different populations.
Most adult Whitethroats Sylvia communis breeding in Europe have a complete or almost complete moult after the breeding season, prior to autumn migration, while populations from Asia ( S. c. icterops, earlier an intermediary form "volgensis" in S. Russia was acknowledged too) either exchange S7 - 9, one or a few rectrices and inner primaries, then suspend and complete in winter quarters, or migrate with old remiges and start moulting very late in East Africa, in December or January. 1y birds are said to have a complete moult by the same time as well (Stresemann & Stresemann 1968) - it is unclear why Svensson 1992 writes "WC (?)" in the case of young icterops. As a consequence eastern populations arrive at breeding-grounds in spring with rather fresh plumage, while European birds - in particular first-time breeders - are fairly worn by that time. In July before the moult starts, the remiges of most European birds are in an extremely poor state. Stresemann & Stresemann wrote about this as early as 1968, (Kenneth Williamson was first with "Identification for Ringers. The Genus Sylvia" containing notes on icterops moult, it appeared in 1964), later on the moult of British Whitethroats has been treated by Pimm 1976. In "Moult in Birds" Ginn & Melville 1983 compile 837 British moult-cards; here the duration of primary moult is said to be appr. 40 days.
The moult of the genus Sylvia tends to be "messy" in general, from Sweden suspended moult has been described in the Barred Warbler Sylvia nisoria, (Hasselquist et al. 1988) and the Garden Warblers Sylvia borin may exchange a few remiges in southern Sweden and then suspend. Lesser Whitethroats Sylvia curruca and Whitethroats often have irregularities in the sequence of secondaries; S6 is sometimes shed together with S7 - 9 ("the tertials"), and even S4 and S5 are in some cases completed before S1 - 3. In other cases S3 - 4 or S4 - 5 or even S4 - 6 are not exchanged in late summer, where this occurs they were probably exchanged together with S7 - 9 in winter quarters and are less worn than the rest of the remiges. So, there seems to be a weak connection between S4 - 6 (3 - 6) and S7 - 9, that disturbs or upsets regularity in the case of Whitethroats. A few moult-cards illustrating this irregularity are presented in full below, the site is Ljunghusen, 55.24 N, 12. 55 E, SW Scania, Sweden:
From this list it is clear, that many Whitethroats do not perform a complete remige moult in Sweden before the autumn migration; 2, 3, 4 and in one case all six secondaries were saved for a later occasion. As a matter of fact this strategy may be very widespread, even a majority strategy. I have old moult-cards, where birds are said to have had score "149" and "150", i.e. where I have taken for granted that all fifteen remiges were exchanged, but it may not have been so in all these cases. It was not until the nineties that I began to look closely at secondaries and discovered the difference between fresh and half-fresh feathers in August (the difference between old and new remiges is impossible to overlook at any time of year, the difference between half-worn and fresh somewhat more difficult to detect; my crucial error was to assume that unshed secondaries were going to be shed later on. But if S3 (worn) is unshed when S2 has grown 95 % it will not be shed. On the other hand it may still be worth while checking S6 and S5, these are shed more independently!). This error on the part of ringers is probably rather common where Whitethroats are involved, and it may occur with other species as well. The termination of moult will still be correctly estimated, even if a bird with 12 fresh remiges is given score 150, but if such points are used in regression analysis together with scores from birds that have actually exchanged fifteen remiges, the duration of individual moult (with 15 remiges involved) will not be correctly estimated.
There are 65 moult-cards at hand: 49 recorded by myself at Ljunghusen, 16 recorded by Ulf Lundwall near Lund and at Löddesnäs, all sites in SW Scania. The distribution of the material is shown in Table I:
| P + S: moult score | n | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 0-30 | 29 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 31-60 | 6 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 61-90 | 2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 91-120 | 6 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 121-150 | 22 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The material has the form of an hourglass. When males aren't singing, Whitethroats with their hidden way of life do not need much "air transport", and when watched in the field it seems as if their flying ability is unimpaired, however large the moult gaps. They are probably capable of increasing the wingbeat frequency. Still, in the mid-phase of moult, they go into hiding, clinging and jumping in thick undergrowth, avoiding to expose themselves in flight. This is the reason why few Whitethroats are caught during this phase. In spite of this deficiency, the material in its present state allows calculations, the results are shown in Figs. 1 and 2, TIME 1 = 15 July. Following Ginn 1975 the regression TIME against MOULT SCORE is used to estimate the average progress of moult in individual birds, the regression MS against TIME to estimate the average stage of moult of the population.
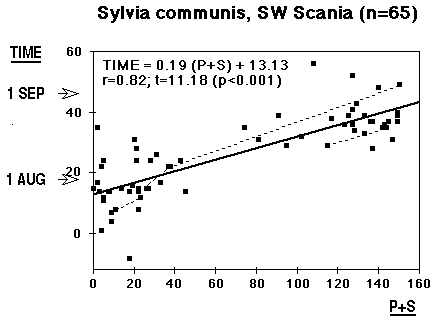
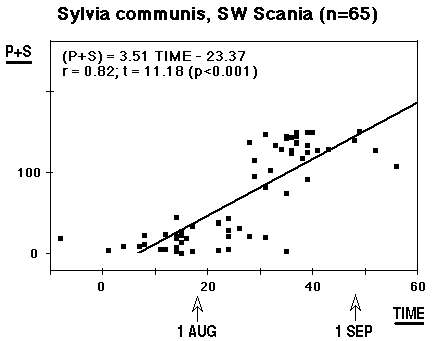
At first sight the result is a little shocking; (as long as the individual cards have not been investigated): individual moult lasts on average 29 days, between 27 July and 24 August, and the population is on average involved in moult for 43 days, with average start 22 July and average termination 2 September. The material looks fine, the correlation coefficient is better than in many other comparable cases (Bearded Tit: 0.69, Willow Warbler: 0.77 - see moult section). The three controls do not contradict the calculated value; they indicate two or three days longer duration - but not ten. Could the passing over of a few secondaries gain that much time (ten days)? Under all circumstances it is not very rewarding to compare with other calculations (or estimates); either the same phenomenon occurs in other areas, too, but has escaped notice there, or the suspension / arrest of secondary moult is restricted to the northern limit of the area, where Whitethroats lay two broods. I think the explanation should be looked for here; in SW Scania Whitethroats often feed begging young as late as mid-August.
In Moult in Birds the duration of individual moult (of British Whitethroats) is estimated at 40 days, but the authors may have been a little generous here; the shaded diagrams with side-lines drawn often conceal more than they reveal. It is noticed (references: Mead & Watmough 1976, Swan & Baillie 1979) that 3 % of all trans-migrants investigated in Iberia and as much as 67 % of all investigated in Crete had one or several old secondaries. Birds of Scandinavian origin occur in Spain, a few birds from eastern Sweden and birds from Finland in Crete. Stresemann notes that S6 is often shed before S5, and finally remarks:"...and we have often noted apparently chaotic leaps." These leaps may not be entirely chaotic when it comes to it; secondaries that were moulted in winter in Africa are retained. A similar leaving out of secondaries is suggested by Hansen 1985 in the case of the Spotted Flycatcher Muscicapa striata, here secondaries exchanged before the autumn migration are probably left untouched in the complete moult in winter quarters.
Why then do eastern Whitethroats migrate in autumn with mainly worn wing + tail, while western Whitethroats do the same thing in spring? Stresemann & Stresemann 1968 point to the fact that the easternmost populations from Asia migrate some 9000 km and winter mainly south of the equator, while western populations seldom migrate more than 5300 km and winter north of the equator. Similar patterns in different populations of Charadrius hiaticula and Acrocephalus arundinaceus are referred to for corroboration of the hypothesis. This sounds attractive enough: it takes longer for the populations flying the longest distance before they are "settled" in a wintering area; Williamson (1964) notes that "in N.E. Africa (Ethiopia, Somalia) the moult may be up to three weeks earlier than farther south (Uganda, Tanzania)". Still, the impression gained from the examples quoted by Williamson is that of an extremely "patchy", opportunist, ad hoc moult where eastern Whitethroats are involved; they use what little resources there are to patch up some sort of complete feather exchange in the breeding area, underway and in the final wintering area. As far as I can see, from general considerations based on knowledge of the variations in precipitation in Africa, the individual moult pattern of Whitethroats belonging to east Asian populations must differ strongly between years.
Distance cannot hold the clue to this problem and I strongly doubt that a conclusive cost-benefit calculation could be construed, showing that it is more advantageous to fly the longer distance with worn remiges in autumn and fresh remiges in spring, and vice versa for the shorter distance. There is ample insect food (qualitative food needed for moult!) for moulting Whitethroats in Western Europe in late summer, but if we look at e.g. Romania or Thrakia in eastern Greece by the same time, the problem to eastern Whitethroats becomes obvious: the countryside is extremely barren and dry and insects are rare. The same applies to the whole of the Central Asian area in July, August and early September. Berries and fruits are abundant by this time of year, these can be used for fattening (followed by migration), but moult can not be based on berries. So, the main cause can not lie in distance, instead feeding conditions and rainfall in both the breeding area and in Africa and in the breeding area must combine to create the moult patterns of different Whitethroat populations. Although the amount and the timing of African rainfall is highly variable, East Africa south of the equator has two distinct rain periods, one around May, one minor round late October. In contrast West Africa has only one major rain period: mainly from May to September (Moreau 1972). And we know, that many wintering Palearctic passerines (Riparia riparia, Phoenicurus phoenicurus, Saxicola rubetra, Sylvia communis) congregate in rather dry parts of the Sahel region, as far to the north as the Niger bend. Given the climatic preconditions, the prospects for moult in the Sahel zone in autumn do not seem very attractive, while there might be better chances (but no guarantee, therefore: "carpe diem") in East Africa. The incidence of secondaries and rectrices exchanged in winter quarters in European populations in turn may vary with precipitation; unexpected winter rain, whenever it occurs, could trigger some "extra" moult.