
|
|
Backsvalans övergång från häckning till flyttning under eftersommaren |
| For readers from other continents: The Sand Martin Riparia riparia breeds on the whole northern hemisphere. Young start to migrate from Scandinavia already in July, the main wintering area is situated in the Sahel zone in Africa. |
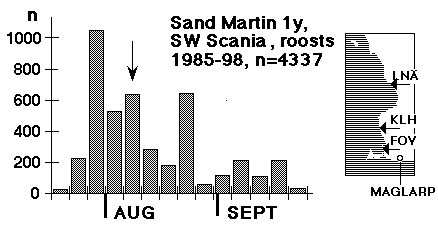
 En enskild fångst visade vad vi hade gått miste om: den 23 augusti 1985 fångades i Foteviken, med åtta nät satta just med sikte på svalorna, 343 unga och 12 gamla backsvalor (andra arter oräknade). Detta tillfälle visade att det fanns en potential, men samtidigt att man är tvungen att akta sig när man går efter svalor - också med en så pass begränsad nätmängd som åtta nät. Endast flyhänta och välutrustade märkare skall ge sig på denna typ av fångst. Efter detta isolerade försök dröjde det ett par år innan det gjordes nya försök, men under åren 1991 - 95 märkte framför allt Peter Olsson, med assistans i fält av Kerstin Norrman och Per Nothagen ungefär 4000 backsvalor i Klagshamn och Foteviken, samtidigt som Christer Persson fortfarande märkte i de närliggande kolonierna (i första hand Maglarp). Här avkastades med en gång rätt mycket kontroller i bägge riktningar, och det är framför allt materialet från dessa fem "insats-år" som ligger till grund för denna uppsats. Den tidsmässiga fördelningen för 5199 ungfåglar, märkta i kolonier mellan Ven och Skurup mellan 1985 och 1998 visas i fig. 1, den tidsmässiga fördelningen för 4337 ungfåglar märkta i sovvassar i Foteviken, Klagshamn och Löddesnäs under samma skede visas i fig. 2, de tre viktiga sovvassarnas samt Maglarps läge framgår av kartskissen bredvid diagrammet i fig. 2.
En enskild fångst visade vad vi hade gått miste om: den 23 augusti 1985 fångades i Foteviken, med åtta nät satta just med sikte på svalorna, 343 unga och 12 gamla backsvalor (andra arter oräknade). Detta tillfälle visade att det fanns en potential, men samtidigt att man är tvungen att akta sig när man går efter svalor - också med en så pass begränsad nätmängd som åtta nät. Endast flyhänta och välutrustade märkare skall ge sig på denna typ av fångst. Efter detta isolerade försök dröjde det ett par år innan det gjordes nya försök, men under åren 1991 - 95 märkte framför allt Peter Olsson, med assistans i fält av Kerstin Norrman och Per Nothagen ungefär 4000 backsvalor i Klagshamn och Foteviken, samtidigt som Christer Persson fortfarande märkte i de närliggande kolonierna (i första hand Maglarp). Här avkastades med en gång rätt mycket kontroller i bägge riktningar, och det är framför allt materialet från dessa fem "insats-år" som ligger till grund för denna uppsats. Den tidsmässiga fördelningen för 5199 ungfåglar, märkta i kolonier mellan Ven och Skurup mellan 1985 och 1998 visas i fig. 1, den tidsmässiga fördelningen för 4337 ungfåglar märkta i sovvassar i Foteviken, Klagshamn och Löddesnäs under samma skede visas i fig. 2, de tre viktiga sovvassarnas samt Maglarps läge framgår av kartskissen bredvid diagrammet i fig. 2.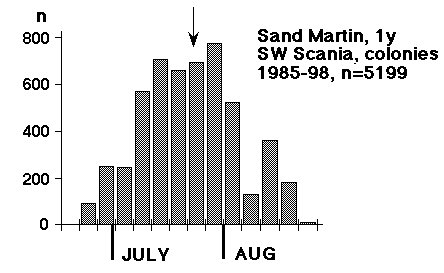
I olika sammanhang, exempelvis hos Emlen & Demong (1975), har man pekat på den uttalade samtidigheten hos häckningen inom enskilda kolonier eller koloniavsnitt; synkroniseringen skall leda till en eller annan fördel: "An individual that fledges either early or at the peak of synchrony will emerge to find a steady stream of other birds traveling to local ephemeral concentrations of food". Detta flitigt traderade påstående (som nog hämtat inspiration från arbeten om kolonihäckande sjöfåglar) måste få en kommentar innan vi kan gå vidare. Det är korrekt att en grupp fåglar, som gemensamt tyr sig till ett väggavsnitt och gräver bohål där, ligger i fas med varandra när det gäller äggläggning, kläckning och ungarnas utflygning. Men redan inom en "koloni" (ett sandtag) finns det för det mesta delkolonier som är tidsmässigt sidoförskjutna (grävande, ruvande och matande fåglar inom hundra meters avstånd), och när man rör sig inom ett större område är det uppenbart att närliggande (inbördes avstånd 2 - 3 kilometer) kolonier sällan är i fas med varandra. De drar sina fåglar från samma pool, vilket leder till att fasförskjutningen kan bli så stor som tio dagar. Mycket hos backsvalan i Nordvästeuropa går med nödvändighet, på grund av det nyckfulla sommarklimatet, ut på riskspridning, fåglarna får under inga omständigheter "lägga alla ägg i en korg". Denna riskspridning, som man i viss mån kan ana sig till redan av fig. 1 - ingen annan art (möjligen med undantag för skäggmesen) slussar ut ungar på det sättet under mer än en månads tid - effektueras dels genom ett utdraget vårsträck, som leder till att många fåglar uppenbarligen infinner sig i kolonin först i början av juni, dels uppkommer den på "naturlig" väg, genom ras och predation. Under en utsträckt period - 2 till 2 1/2 månader - möter fåglarna varje sådan motgång genom att bilda en ny grupp och gå in i häckning på nytt. Det råder ingen övergripande synkroni, bara ett stort tumult, och det finns alltid gamla, matande fåglar till hands för att peka ut exempelvis "ephemeral concentrations of food".
I flerkullsfallet rullar häckningen vidare så att de flygga ungarna "vid någon tidpunkt" lämnar kolonin, i sällskap med de hanar som är deras fäder, detta dränerar fortlöpande kolonins hanbestånd. Samtidigt parar honan om sig med en ny hane, om en sådan finns tillgänglig. Därefter går hela häckningsproceduren ett nytt varv. Denna uppsats handlar om flerkullsbacksvalor och en aspekt på deras fenologi: tidsschemat för bortflyttningen. Som framgått av det ovanstående finns det två problem som får en eller annan "lösning" inom kolonins ram, det första ser ut så här:
Detta upplägg har ett syfte som är nämnt redan i introduktionen: jag vill i utgångspunkten undvika adaptiva efterhandskonstruktioner, tona ned vikten av selektivt gynnade mainstream-lösningar på olika valsituationer som löses inom ramen för den sociala strukturen. (Storheter som kullstorlek, fettnivåer, allt som bygger på rent fysiologiska instruktioner och utspelar sig mer eller mindre på det fysiologiska planet, faller utanför detta resonemang). Sociala valsituationer som inbegriper interaktion mellan individer är oåtkomliga för "instruktion", de löses ad hoc, på stället, denna "lösning" är det allt avgörande, och ett flertal olika alternativ kan integreras och fås att fungera inom helhetens ram. "The adaptive significance" hos synkronisering, icke-synkronisering, tidig flyttning, sen flyttning är, när alla led kommer med i ekvationen (och: inom vissa gränser) - noll, zero. Det avgörande är hur populationen opererar med sina parametrar, medan de fördelar som den själviska individen eventuellt kan tillskansa sig skingras som agnar för vinden. Nu till det fältmaterial, som belyser fåglarnas övergång från kolonin till sovvassarna och den efterföljande bortflyttningen.
De tidiga sommarrörelserna, eller rentav den tidiga sommarbortflyttningen hos en handfull arter (adulta rörsångare, sävsångare, kärrsångare, härmsångare, grönsångare och - unga backsvalor) är inte fullt belyst i det inledande skedet; av fågelskyddsskäl drar man sig ofta för att fånga i vassarna före ca 20 juli (Flommen, Klagshamn), och i Foteviken kan det finnas sena kullar av ex. skärfläcka som gör att ringmärkaren gärna avstår från större fångstinsatser fram till förste augusti. Men det är alldeles uppenbart att de ovannämnda arterna "trycker på" lite vid den skånska sydkusten redan från omkring 15 juli, och åtminstone gamla härmsångare och grönsångare (kanske fåglar som förlorat kullen) kan nog börja flytta så tidigt som 10 juli. När man väl har vant sig vid denna tanke är det lättare att acceptera att backsvalans flyttning också inleds någon gång i juli, och att vi rätt säkert saknar data från den inledande fasen också när det gäller denna art. Som upptakt till bortflyttningen samlas fåglarna för gemensam övernattning i "sovvassar", som sådana tjänar bl.a. Flommen i Skanör, kustvassarna från Foteviken och norrut och grova vassar i dammar (Lockarp, Torreberga) och sjöar (Krankesjön, Yddingen etc.) inåt land. Det finns alltid en viss tradition knuten till sovvassarna; de vuxna fåglarna känner till deras lägen, och i kolonier med märkbar predation sover hanarna överhuvud taget inte i bona, utan i en närliggande sovvass.
I fig. 3 redovisas alla juvenila backsvalor som märkts i kolonier från Ven i norr till Maglarp i söder (ett dussin kolonier är involverade, men merparten av materialet kommer från Maglarp), och sedermera kontrollerats i sovvassar, samt alla fåglar som kontrollerats två gånger samma säsong i sovvassar. I genomsnitt 8 � 5 dagar efter att den märkts i kolonin har en ungfågel återfångats ute i sovvassen, två fåglar har rentav tagits där på kvällen samma dag som de gryningsmärktes i kolonin. De första fåglar som överhuvud taget kontrollerats i sovvass (den 23 juli) har varit märkta i koloni den 17 juli. Medianen för ungfågelsmärkningarna i kolonierna faller på den 23 juli, vilket innebär att i runt tal 40 % av märkvolymen inte avkastat en enda kontroll i sovvassarna. Vi kan alltså rätt säkert utgå från att tidiga ungfåglar är kapabla att lämna kolonin och övergå till att sova utanför denna redan i första dekaden av juli, och det verkar rimligt att anta att de första hanarna flyttar söderut vid samma tid. Lika intressant är att ungfåglarna inte heller tycks uppehålla sig länge i sovvassarna: sex fåglar har kontrollerats i dessa i snitt 4,5 dagar efter märkningen, och ingen ungfågel, vare sig från koloni eller från sovvass, har kontrollerats där två gånger. Fig. 3 illustrerar brådska, högt tempo, och kanske ett tempo som är mest tillspetsat under första tiodagarsperioden i augusti.
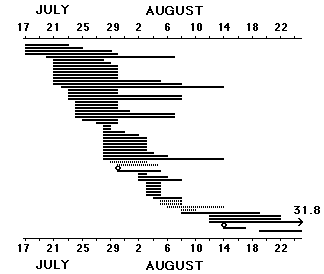
Till detta diagram kan läggas en 1k-fågel, som märktes på Flommen, Falsterbo den 22.7.88 och kontrollerades i Maglarp 89, samt en 1k från Klagshamn 16.9.93, som kontrollerades i Maglarp 95. Vi har alltså lokala ungfåglar i de sydvästskånska sovvassarna åtminstone under intervallet 22.7 - 16.9. Nu har inte alla fåglar lika brått att lämna kolonimiljön (med boet eller bogången som välkommet skydd mot nattliga regn); en icke försumbar mängd stannar kvar rätt länge i kolonin. Ett urval (en stor del av dessa data ligger fortfarande oanalyserade i anteckningsböckerna) av sådana kontroller visas i fig. 4. I snitt har de 31 fåglarna uppehållit sig i kolonin ytterligare 13 � 5 dagar efter märkningen. Tempot är alltså inte lika högt här som i sovvassarna, men det kan bero på att materialet omfattar yngre, mer outvecklade fåglar, eller att det kommer från regniga, för fåglarna oynnsamma år. Vi kan inte heller utesluta en viss störningseffekt, som påskyndar utvandringen till sovvassarna. (Dessa frågor lämnas tills vidare öppna).

Ett tredje alternativ är att ungarna ansluter sig till en annan koloni och utnyttjar dess lediga bogångar för övernattning. Samtliga kontroller i denna kategori är redovisade i tabell I, det genomsnittliga uppehållet var 13 � 8 dagar, alltså samma varaktighet som för de längsta uppehållen i födelsekolonin.
| Märkdatum | Distans (km), kurs | Intervall (dagar) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 25.6 | 18 NW | 26 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 25.6 | 28 W | 9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 27.6 | 30 ENE | 14 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.7 | 4 SE | 13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.7 | 13 SSW | 5 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.7 | 48 SSW | 16 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.7 | 23 NE | 10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.7 | 4 NW | 26 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.7 | 12 SSW | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 14.7 | 7 ESE | 4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 18.7 | 0.5 SW | 1 timme | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20.7 | 12 SSW | 11 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20.7 | 3.5 W | 15 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kön, antal | Intervall | Median | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| hanar (märkn.): 134 | 20.7-29.8 | 30 juli | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| honor (märkn.): 152 | 20.7-12.9 | 5 augusti | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| okön. (märkn.): 117 | 25.7-17.9 | 19 augusti | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| hanar (kontr..): 28 | 20.7-19.8 | 31 juli | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| honor (kontr..): 23 | 23.7-25.8 | 6 augusti | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Till detta skall fogas en kontroll, som visar att inte ens alla adulta hanar har gjort ett slutgiltigt val när de besöker sovvassarna:
Just 1995 födde en del honor i Maglarp rätt säkert upp tre kullar, och hanarna var efterfrågade mot slutet av säsongen. Det är mycket tänkbart att denna fågel beledsagade ungar ur andrakullen till Foteviken och sedan vände tillbaka för att bidra till uppfödningen av en tredje kull. (Men en annan möjlighet är att den regelbundet sov i Foteviken för att undvika grävlingpredationen i Maglarp).
Själva "samlingen" i sovvassarna tycks gå ut på att få ihop en tillräcklig flockstorlek (säg: tusen individer eller mer), därav den egendomligt pulserande karaktär hos uppträdandet, som alltid noteras i fält, och som också framgår av Fig. 1. I histogrammet går det att urskilja en kulmination för en "förstakull" kort före eller kring 1 augusti och kulminationen för en "andrakull" kring 20 augusti, därtill finns den vänsterförskjutning, som är utmärkande för långdistansflyttare (Dorka 1966). Men man kommer i backsvalans fall aldrig att få det jämna, kontinuerliga diagram, som är utmärkande för arter som exempelvis gulärla och lövsångare. En lugn och vacker kväll kan det sova tretusen backsvalor i Foteviken, nästa kväll, som är lika fin, bara tvåhundra, då förstår man att en kontingent gett sig iväg, samtidigt som uppbyggnaden av nästa flock redan pågår. (Men ibland handlar det bara om sovvassbyte på grund av exempelvis närgångna sparvhökar). Sist och slutligen blir det ändå den tid det tar för att bygga upp en flock, som avgör hur länge en individ uppehåller sig i sovvassen; det är rimligt att anta att detta uppehåll blir kortast kring 1 augusti, när genomströmningen av fåglar är störst. Här drar jag streck; materialen är för små för att pressas till utsaga på denna punkt.
1. Tempot
Det finns en populär och flitigt traderad bild av svalorna som "samlar sig" inför höstflyttningen och så småningom drar iväg mot Södern i samlad tropp. Denna bild svarar dåligt mot verkligheten; "borttransporten" av sydskånska ungfåglar inleds i själva verket mycket tidigt på säsongen (15 juli kan få stå som provisoriskt startdatum, men vi skall nog ner till 5 juli), och pågår sedan i högt tempo säsongen ut. Skillnaden mellan medianerna i fig. 1 och fig. 2 ger ett närmevärde på hur lång tid den flyttande ungfågeln varit flygg: cirka 17 dagar. Samma höga tempo förekommer också längre söderut; enligt Mead & Harrison (1979b) gick den snabbaste brittiska ungfågeln från Fife, Skottland (märkt 24 juli) till Bordeaux, Frankrike på fyra dagar - en sträcka på 1242 km! Och precis som i Skåne var det fastställda medeluppehållet i den stora sovvassen vid Chichester (Sussex) mycket kortvarigt: 7 resp. 8 dagar för ungfåglar och adulta (Mead & Harrison 1979a). (Ett liknande tempo tycks utmärka den unga kärrsångaren; av duntillväxten under konturfjädrarna att döma kan en del individer inte ha varit ur boet mer än två veckor, när de dyker upp vid skånska sydkusten i sena juli. B. Petersson, muntl.). De första gamla hanarna av backsvala lämnar också landet kring mitten av juli, medan kolonilivet mestadels tar honorna i anspråk till ett stycke in i augusti. Redan kring 1 september har praktiskt taget alla häckfåglar (utom en mindre mängd som vissa år knogar med en tredjekull eller en sen ersättningskull) lämnat Skåne. De backsvalor, unga såväl som gamla, som fortsätter att utnyttja sovvassarna i september tycks i huvudsak vara av nordligt ursprung. (Två uppsatser: om de gamla fåglarnas uppehållstid i kolonin och om nordliga backsvalors sträck kommer längre fram).
2. Alternativen
Flerkullsbacksvalornas häckning framstår som om den ägde rum i en box med snävt satta, orubbliga väggar: allt skall göras färdigt under tre sommarmånader, och som en extra besvärande omständighet är en svensk juni ofta för kall för svalor, fåglarna hankar sig fram och kommer inte igång ordentligt, ungarna växer på sparlåga inne i bona. Då blir det extra viktigt att lägga på ett kol när födan äntligen finns där och unguppfödningen löper. Backsvalans speciella häckningssystem (Cowley 1983) är inriktat på att uppnå maximalt utbyte i form av flygga ungar under detta skede. Men därutöver finns en andra förutsättning: en sorts robusthet hos ungarna, en förmåga hos dessa att ta vara på sig. Detta utnyttjas så att de slussas in på en mellanstation: sovvassarna, den egna kolonin eller andra kolonier, där de kan tillgodogöra sig information från de vuxna fåglarna. Rätt fort får de däremot klara sig utan individuell tillsyn från de vuxnas sida. I samtliga fall, hanarnas, honornas och ungfåglarnas, handlar det om att följa, en sorts associerande process, där hanarna intar en central position:

3. Ungfågeln skall "följa"
Hanen har know-how: han vet när det finns tillgängliga honor i kolonin (i kraft av förföljelseflykten), han kan föra mycket små ungar från kolonin till ett skyddat läge eller en annan koloni och fortsätta att mata dem där, och han kan styra ut välutvecklade ungar mot sovvassarna. På detta sätt intar han en nyckelställning som informatör. Samtidigt har ungfågeln fortfarande ett val, den kan skaffa sig information av någon annan än den kullförande fadern. Om den inte är mogen att lämna kolonins skyddade miljö kan den försöka reda sig en tid på egen hand inom en kolonis ram, och den kan göra samma sak i en främmande koloni. I bägge fallen heter lösningen "följa John" (följa förebilden), den grundläggande processen är associativ. Ändamålet i form av ex. predationsskydd rör aldrig vid denna fundamentala, formbildande eller dynamikskapande process (den måste vara grundläggande pga ungfåglarnas totala oerfarenhet; den går i spåren på embryonalutvecklingen), formbildningen är icke-intentionell och alltid möjlig att upprätta eller rekonstruera på denna nivå. Slutresultatet av den dynamiska interaktionen för ungfåglarnas del blir en slutgiltig avresa med kurs på vinterkvarteren.
På detta sätt kan backsvalor till och med överbrygga eller kompensera för katastrofer av det slag som drabbade dem 1968 och 1980. Samtidigt är det uppenbart att resultaten av den dynamiska interaktionen, skapandet av form i populationen - rent hypotetiskt - kan instrumentaliseras och mobiliseras i ett diakroniskt (historiskt eller evolutionärt) perspektiv, exempelvis som predationsskydd. (Vuxna fåglar varnar, sluter sig samman och tar höjd inför sparvhök och lärkfalk; ungfåglarna tar efter detta beteende redan första dagen i det fria). Men när allt kommer omkring är de flesta av dessa "anpassningar" blott skenbara, de är inget annat än resultat på platsen av formbildningsprocessen, och denna kan röras bara av revolution, inte evolution. (Samma slag av process omfattar naturligtvis också andra arter: bofinkar, trastsångare eller halsbandsflugsnappare - det är bara att vända på kikaren och "packa ihop" en revirfördelad population så ser man den speciella, artkarakteristiska struktureringen. Backsvalan däremot inbjuder direkt till detta synsätt genom sin kolonialitet, genom det faktum att en individ alltid har en annan individ i backspegeln. Hon är i inget sammanhang en individuell atom, utan alltid del i en struktur, en väv, ett gitter.)
4. En modell för den dynamiska interaktionen mellan backsvalor före bortflyttningen
När jag vill uppställa en modell för det dynamiska mönster, som skildrats ovan, tillgriper jag den elementära katastrofteori, som från och med augusti 01 finns tillgänglig i teorimappen. Detta teoriavsnitt måste läsas (och begripas) innan fortsättningen blir möjlig att ta till sig. Nödvändig förkunskap är ett hum om funktionsteori (man skall veta vad derivering går ut på och helst ha stött på gradientfunktionen), och kanske någon känsla för tre- och fyrdimensionell geometri; finns inte detta till hands kan man hoppa över den närmaste fortsättningen. För mer djupgående inträngande i matematiken krävs minst gymnasiets mängdteori, och helst de första kurserna på universitetsnivå (man har exempelvis mycket gratis om man begriper lineära transformationer, kvadratiska och kubiska former). Men man kan också reda sig en bra bit med lite självstudier i ämnet!
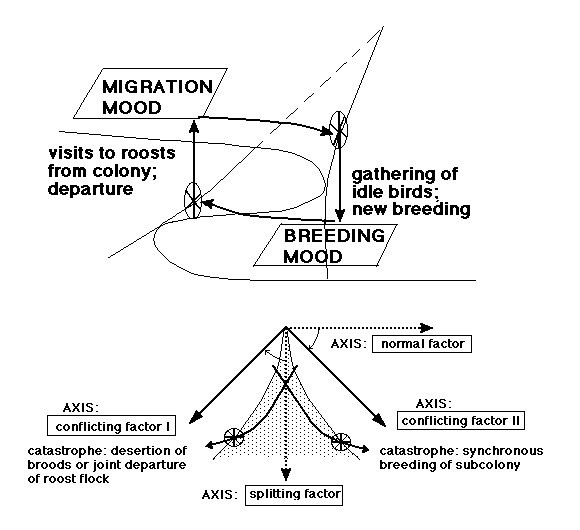
Bland backsvalor som uppehåller sig på nordliga latituder på sommaren finns två motstridiga behov: dels behovet att föda upp så mycket avkomma som möjligt, dels behovet att flytta bort vid en tidpunkt som är optimal för populationen (inte individen). Dessa behov kan inte regleras med hjälp av en fix kalender, lösningen måste i stället heta en dynamisk interaktion, där information tas fram och överförs till de deltagande fåglarna. Här har vi ett typiskt bimodalt mönster (flytta/häcka) med plötsliga språng (hastig avfärd, sena häckningsförsök) och hysteres (retur till kolonin från sovvassarna, övergivande av sena kullar - eller, viktigt, övergivande vid vilken tidpunkt som helst på säsongen), som inbjuder till ett modellerande med hjälp av "cusp"-katastrofen. Samtidighet i häckningen och häckningssammanbrott är två sidor av samma mynt (även om de inte uppträder med samma frekvens), detta är en oerhört viktig insikt. Och en annan viktig insikt är: den primära, topologiska nivån ger i backsvalans fall företräde åt en sorts gruppselektion, individselektionen får sitt spelrum beskuret. (Jag vet inte hur långt detta kan generaliseras, men min övertygelse är att all dynamisk interaktion - dvs. också växelspelet mellan revirhävdande arter - på ett eller annat sätt kan och bör föras in under katastrofteorins tak. Två-tre års fältiakttagelser av skärfläckan i Skanör har i vart fall demonstrerat för mig, att både synkronisering, sammanbrott och individuella "avvikelser" hos denna art lyder under samma katastrof-dynamik som i backsvalans fall, och jag har en stark känsla av att småtärnan skall föras till samma läger. Synkroniseringen är aldrig total, utan omfattar delkolonier, häckningssammanbrottet är ett faktum långt innan predation eller väder och vind förstört alla kullar, och enstaka par går alltid sin egen väg, bryter mot trenden. Fjällabben kan vara en viktig länk mellan revirhävdare och kolonihäckare; den är territoriell, men varje par vet var grannparen finns, här finns en myckenhet av wynne-edwardskt "display", häckningen är synkroniserad, sammanbrotten likaså, och "tokiga" avvikare stöter man på med jämna mellanrum).
Hos backsvalan står ett plan (hos det dynamiska "vecket") för förstärkt flyttningsbeteende, det andra för förstärkt häckningsinsats, och sprången (katastroferna) från ena planet till det andra styrs av två kontrollparametrar, som inledningsvis består av en normalfaktor (jag föreslår: ett vägt medelvärde av antalet honor + födotillgången), och en "splitting"-faktor: häckningsframgången. Det finns återkopplingar: efterfråga på hanar sänder dem tillbaka från sovvassarna till kolonierna, medan brist på honor i förtid sänder hanar till sovvassarna. Återkopplingarna är försöksvisa, långsammare, de sker som ett läckage, den "stora" samtidigheten lyser med sin frånvaro; om tidsskalan för katastrofen är en dag, är den två veckor för återkopplingen. Här finns det individuella spelrummet, en mycket liten grupp kan alltid bryta mot majoritetsbesluten. Efterhand som säsongen framskrider vrids startsystemet medurs till ett system med två motstridiga kontrollfaktorer (eller perspektivet växlas genom en transformation: normal + splitting = control II, normal - splitting = control I): här måste den ultimativa faktorn (kontrollfaktor I) i ena fallet vara dagslängden, dess proximativa yttring fysiologisk förändring (indikerad av begynnande kroppsfjäderruggning), medan kontrollfaktor II fortfarande handlar om tillgången på häckningsbenägna honor och födotillgången. Härigenom reduceras både antalet honor och häckningsutfallet, till slut är antalet häckningsdugliga honor noll och en slutgiltig katastrof sänder de resterande fåglarna till vinterkvarteret. Hela anordningen framgår av Fig. 6:
5. Adaptionstänkandets invändningar
Invändningen mot förslaget att det ena valet kan vara så gott som det andra kan lyda: Varför går de första ungfåglarna då iväg så tidigt, och varför flyttar vissa gamla fåglar tidigt, i stället för att satsa på häckning ännu en tid? Någon form av fördel måste vara förbundnen med den tidiga avfärden och det höga sträcktempot. Tidigt kläckta ungfåglar har högre överlevnad än senare kläckta (Persson 1987), det är dessutom frestande att föreslå att de genom sin avfärd "skapar utrymme" för boungar och äggkullar genom att inte tulla på kolonins födoförråd. I SV-Skåne faller denna invändning till marken; alla sovvassar ligger inom den dagliga aktionsradien (10-15 kilometer) för adulta fåglar från ett flertal stora kolonier.
De vuxna fåglar som ger sig tidigt av kommer i sin tur först in i en ruggning som varar i 4 1/2 månad (Ginn & Melville 1983), och naturligtvis gynnas även tidigt flyttande årsungar på samma sätt. Det kan dessutom vara nyttigt att ta sig över Medelhavet respektiva östra delen av Sahara innan eleonorafalkens och sotfalkens häckning kulminerar (eleonorafalken ca 25 augusti till 25 oktober, Stresemann 1968, Walter 1968), backsvalor ingår i dieten hos bägge arterna. I såväl de gamlas som de ungas fall har det adaptiva tänkandet ett bud på saken, men är det positiva utfallet i något sammanhang kristallklart, evident - är "anpassningen" överhuvud taget så välbetänkt? De gamla fåglarna och ungfåglar som ligger långt framme på hösten återvänder först på våren (referens för de gamla fåglarnas del: Mead & Harrison 1979b) - och får sin förstakull mer eller mindre spolierad om juni är kall och regnig. Dessa år är de eftersläntrande förstagångshäckarnas ungar de första att bli flygga (10 - 15 juli). Vilket vill säga: de fåglar som i minst grad demonstrerat sin "fitness" drar högsta vinsten i häckningen... Mead & Harrison (1979b) har en kommentar också här (i förbigående drabbar den tesen om backsvalans långt drivna synkronisation): The later arrival of young birds must mean that they will also commence breeding later. This may explain the continuing succession of nesting attempts that seem to be taking place at any large colony throughout the breeding season, and also the establishment of new colonies in late May (or even early June) if a suitable site is made available...
På samma sätt är det med ruggningen: visa år sträcker sig eftersläpande verkningar av förra säsongens torka från Sahel upp till Nordafrika, då är det besvärligt att anlända i september och börja rugga; det var en sådan initial torka som knäckte backsvalorna hösten 1968. Andra år faller regnen rikligt och tidigt, då räcker insekterna för alla ruggande övervintrare. Slutligen kan torkan göra sig gällande först på våren och drabba såväl ruggningens slutfas som fettanlagringen inför sträcket. Jag har svårt att föreställa mig hur positiva tendenser skall kunna gynnas av selektiva mekanismer under sådana förhållanden. Det ena årets "val" motverkas av det efterföljande årets, den föreslagna selektionens och den resulterande adaptionens trovärdighet pulvriseras i rask följd av en variabel och nyckfull omvärld. Budskapet för backsvalans del lyder: undvik reglementerande anpassning. I stället är riskspridning och variation A och O för varje art som är inte är alldeles förstenad, fastlåst i sitt ekoschema. (Många arter har ju mindre handlingsutrymme än backsvalan, som får sägas vara välutrustad, lyckligt lottad). Denna riskspridning garanteras av den formbildande processen, som inbegriper en viss fördröjning för en del individer.
Naturligtvis släpper darwinismen inte taget: då måste denna riskspridning, detta genomsyrande ad utrumque paratus vara av "adaptiv" natur. Men är det inte en onödig omväg (och en utspädning av adaptionen till intighet): att föra in evolutionärt modifierade individer, för att med fördröjning hantera problem, som faktiskt redan har lösts direkt av individerna på platsen, i deras interaktion? Selektionens potential framstår i många sammanhang som upphaussad, det kan ställas i tvivel huruvida processen, sådan den utmålas i teorin, någonsin förmår leverera den önskade produkten i rätt tid. Och om leveransen sker, har en annan, kooperativ, interaktiv process redan gjort jobbet.
6. En preliminär sammanfattning:
Jag vill alltså tolka de data som framlagts här på följande sätt, i följande språkdräkt: sovvassarna (de primära) ligger inom koloniernas dagliga aktionsradie. Den första hane, som tillsammans med ett antal ungfåglar väljer att övernatta där, sätter igång det skred som bortflyttningen innebär. Den lilla pionjärgruppen kommer att attrahera andra grupper, som är på vippen att ta samma steg, och samtidigt kommer bortfallet av hanar i kolonin att märkas som en förändring hos signalnivån i kolonin: en hona som tar sig en sväng i luften får inte längre en svans på fyra hanar, utan plötsligt bara tre eller två. I sovvassarna bidrar kvällarnas flygexercis (flockarna bildar täta, manövrerande moln) och intensiva signalerande säkert till att försätta de berörda fåglarna i flyttningstämning: en stor flock håller sällan på med den här typen av exercis mer än en vecka, sedan är merparten av fåglarna borta - samtidigt som nästa flock redan är på väg att skapas. Resonemanget finner stöd i fig. 3; det handlar aldrig om ett längre uppehåll i sovvassen, utan om en sorts inpiskning inför bortsträcket. Den fågel som kommit så långt är förlorad för häckningen - det gäller avgjort för honorna men säkert också för de flesta av hanarna. Samtidigt kan och bör vi inte blunda för en rörelse i motsatt riktning; om inte häcknings-"uppgiften" hade sin dragningskraft, skulle vi aldrig se septemberhäckningar hos backsvalan i Skåne.
För den unga backsvalans del gäller i alla sammanhang en enkel maxim: anslut till andra backsvalor (med ett kärninnehåll av information, buren av gamla fåglar), och låt det resulterande dynamiska systemet av motstridiga målsättningar med hjälp av elementära katastrofer bestämma tidpunkten för bortflyttning, fortsatt häckning, övervintring etc. Och det finns mer att hämta i detta; allt: kontur, struktur, rörelse hos förtätning och utglesning (det vill säga resulterande dispersion och olika typer av "patterns"), bestäms i sin tur av en enkel formbildande process för vilken varje uttalat ändamål är en främmande, genom mänskligt tänkande införd tillsats. Initialvärden kan vara givna på ärftlig väg, omgivningens inflytanden finns där i form av variabla parametrar, som påverkar processen, men denna i sin tur är robust, fundamentalt okänslig, kan svälja stora variationer - och den står ensam för dynamiken. Omvänt kommer den inte att få ett exakt likadant utfall om den upprepas två gånger under identiskt lika yttre betingelser; efter en tid kommer de gamla fåglarna i två parallellt förlöpande proceser med samma utgångsvärden att bära olika information. Därmed skådar vi "evolutionen" i vitögat, min grundhypotes är att det enda som selekteras på den sociala nivån i backsvalans fall är individernas förmåga att associera sig till andra individer, att bidra till att skapa form eller delta i, bidra till den dynamiska interaktionen i populationen. Allt annat i detta sammanhang, alla skenbara "anpassningar" är blinda resultat av den formbildande processen. Jag vill gå så långt som att hävda: efter en tid kommer varje selekterad tendens att bli ett påhäng, en belastning, som den dynamiska interaktionen, formbildningen tvingas neutralisera långt innan samma tendens blivit "motselekterad" - och formprocessen är kapabel till sådan övergripande korrigering.