Hos många småfåglar ruggar de gamla i skarven mellan häckning och flyttning. När ruggningen är överstökad är de färdiga att flytta, och de gör det ofta utan uppskov (om de inte är stannfåglar). Detta är standardfallet, kring det grupperar sig de övriga strategierna. Ofta är tiden för knappt utmätt för att det skall gå att genomföra fjäderbytet i häckningsområdet, då ruggar fåglarna i stället på en mellanstation eller i vinterområdet. Som en upptakt byts ibland ett par pennor före bortflyttningen (exempel: pilgrimsfalk, kärrsnäppa, höksångare), då talar man om uppskjuten ruggning (eng.: suspended moult). Andra arter flyger söderut på gammal vinge och ruggar på en mellanstation, exempelvis grönbenan. Födotillgången spelar naturligtvis också en roll, ruggningen är ett energikrävande förlopp; många invasionsfåglar avbryter ruggningen (eng.: arrested moult) under år då deras stapelfödor tryter (sidensvans, nötkråka, större hackspett). Större arter bär ofta flera fjädergenerationer eller serier (exempel: fiskgjusen, tärnorna), vilket leder till att man får flera, överlagrade ruggningsförlopp. Bakom alla dessa mönster skymtar en komplicerad kalkyl, där naturen under årtusenden har mätt kostnaden för olika ruggningsstrategier mot deras nytta för en viss population. Ruggningsförloppet är tydligt optimerat, det kompromissar sig fram mellan häckning och flyttning, och sneglar hela tiden åt födotillgången. En central uppgift för studiet av ruggningen är att kartlägga det exakta förloppet med alla dess anpassningar och relatera det till födotillgången och de nödvändiga uppgifterna i årscykeln: reproduktion, existens under vinterhalvåret.
VAD REGISTRERAR MAN?
Det tar lång tid att undersöka en ruggande fågel, och också denna undersökning bör optimeras. Av praktiska skäl brukar man tillämpa ett standardiserat förfarande: tillståndet hos de bärande pennorna i stjärt och vinge och hos bestämda "fält" av dun eller konturfjädrar registreras enligt en viss, i förväg överenskommen kod. Vill man hushålla ännu mer med tiden nöjer man sig med att undersöka flygpennorna på ena vingen, alla andra fjädrar brukar normalt hinna bytas under den tid som går från det den första vingpennan fällts tills den sista är utvuxen. Denna undersökning är ofta ett normalt - rentav ett nödvändigt - led i åldersbestämningen; de flesta ungfåglar byter inte pennorna under sin första höst (skäggmesen är ett undantag på småfågelssidan). Hos ungfåglarna brukar man därför nöja sig med att registrera tillväxten av dun på vissa partier av kroppen (som är dunlösa under de första veckorna) och bytet av konturfjädrar till första vinterdräkten.
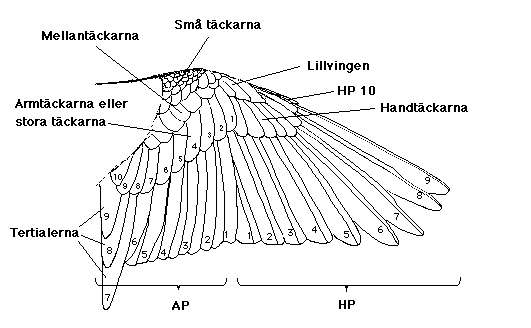
Bytet av ving- och stjärtpennor startar alltid från ett bestämt fokus (eller flera foci). Pennorna numreras utgående från dessa startpunkter, och det första man skall lära sig när man undersöker ruggning är var handpennor och armpennor möts i vingen. Från detta fokus (som brukar betecknas F1) numreras handpennorna utåt (1 - 10), armpennorna inåt (9 hos tättingar, 15-20 hos vadare, tärnor och alkor; jfr Fig. 1). De tre innersta armpennorna kallas tertialer och fälls utgående från ett eget fokus, hos småfåglar ofta först av alla pennor. Innanför tertialerna ligger hos vadarna skapularerna, som ibland kan förtjäna att registreras. En småfågelstjärt (men också en rovfågelstjärt) har 12 pennor, för det mesta fälls det centrala paret först (och denna penna är alltså nummer 1).
Fig. 1. Vingen hos en trädpiplärka. Handpennorna numreras utåt 1-9(10), armpennorna inåt 1-6(9). Någon av A7-9 fälls först, sedan H1. A1 fälls ungefär när handpennorna är till hälften utbytta.
Den europeiska standarden för registrering av växande pennor finns redovisad exempelvis i Ginn/Melvilles "Moult in Birds" (BTO Guide 19). Vi har själva avstått från att bruka denna skala (som urskiljer sex stadier, från 0 till 5), eftersom många har en tendens att glömma bort dess kriterier när de sitter över fågeln. I stället anger vi växande pennors längd i tiondelar av full längd (gammal penna=0, fälld penna=1, kort spole=2, utvuxen penna=10), detta kan sedan lätt räknas om till den allmänna EURING-koden.
HUR VI KOMMER ATT REDOVISA RUGGNINGSDATA
Vi kommer gradvis att arbeta fram en enhetlig form för redovisning av ruggningsdata, en form som dels gör det lätt att plocka ut uppgifter, som samlats på hög i en databas, dels underlättar jämförelsen mellan arter och populationer. I inledningsskedet koncentrerar vi oss på att bearbeta och göra tillgängligt; litteraturutblickarna, kringsynen får vänta till ett senare skede. Den preliminära standard som upprättats i de två första bidragen (skäggmes, lövsångare) ser ut så här:
Där det bara finns ett fåtal undersökta individer, och vi av något skäl vill diskutera detta material, ger vi alla relevanta data, men ofta med inskränkning till ving- och stjärtpennor.
För att man skall kunna bedöma materialets halt gör vi en uppdelning i fem lika stora delar och anger hur många procent av det samlade materialet som faller på vart och ett av intervallen. I det enklaste fallet redovisas dessa värden i formen: IF10-22-31-22-15, utan decimaler.
För att underlätta för utländska läsare (som redan hittat fram till vår sajt) är all diagramtext skriven på engelska. TIME = TID, P (Primary) = HP (Handpenna); S (Secondary) = AP (Armpenna).
Där vi bedömer att vi har tillräckligt med data för att räkna, lägger vi fram materialet som två regressionslinjer: TIME=f(P+S) eller bara TIME=f(P), samt P+S (eller bara: P) = f(TIME). Det första sättet ger ett mått på den enskilda fågelns ruggningshastighet (och linjens fotpunkt anger den genomsnittliga ruggningsstarten), det andra ett mått på det ruggningsstadium, som populationen genomsnittligt befinner sig i vid en viss tidpunkt. I alla sammanhang anges korrelationskoefficienten r i diagrammet. Vidare ges signifikansen t (med n-2 frihetsgrader) för regressionskoefficienten (tangens för linjens lutningsvinkel). Detta görs för att materialet skall bli mer genomskinligt för den statistikbevandrade läsaren. t ger ett mått på sannolikheten att det råder ett lineärt förhållande mellan x- och y-variablerna (vilket det i och för sig nästan undantagslöst gör när man räknar på ruggningsdata; får man ingen regression har man för lite data). Dessa värden betecknas r och t och redovisas med två decimaler i formen r = 0.99, t = 4.19.
I de flesta fall nöjer vi oss med att addera ruggningstillståndet för 9 handpennor och 6 armpennor (den yttersta, förkrympta tionde handpennan och de tre tertialerna har alltså försummas); en färdigruggad vinge (vilket mestadels är detsamma som en färdigruggad fågel) får då "poängen" 150. Det hela har här presenteras i två diagram, behändigare är kanske att arbeta med ett enda, där bägge regressionslinjerna ritats in och materialets yttergränser markerats med streckade linjer (då får det göras större). Under diagrammet följer en kort, kommenterande text - vi skall försöka att hålla den minimal!
Material från olika områden kommer inte att blandas; ett bra material omfattar minst 100 individer från ett område, som inte spänner över mer än cirka 50 kilometer, eller cirka en halv breddgrad.
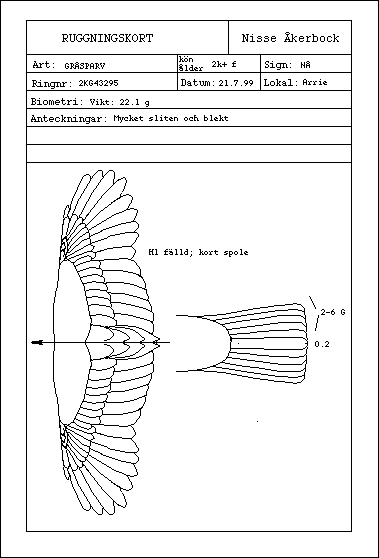
Fig. 2. Mall för ett "ruggningskort", som sedan 80-talet använts mycket av svenska ringmärkare. Man kan ha kortet med ut i fält, men man kan också nöja sig med att fylla i det efter ett avslutat märkpass.
HUR SER UPPGIFTEN UT FÖR SVERIGES DEL TILL ATT BÖRJA MED?
Sverige är ett långsträckt land, spänner över 13 breddgrader och lite till. När man får ruggande fåglar från Lappland i sin hand är det förvånande att se hur lite de skiljer sig från sina skånska artfränder vid samma tidpunkt. Om det råder fjorton dagars skillnad mellan Skåne och Lappland när det gäller äggläggningen, så kan skillnaden på ruggningssidan krympa genom att lapplandsfåglarna relativt sett inleder sin ruggning tidigare och har en snabbare fjädertillväxt på grund av att de kan äta i stort sett dygnet runt. Ett extremt exempel är den nordliga gulärlan, som börjar passera färdigruggad över Skåne samtidigt som lokala flava-fåglar fortfarande har en eller två veckor kvar av sin ruggning. Detta skulle vi vilja ta fram mycket data på: långa serier från norr till söder, som visar hur ruggningsförloppet "kompakteras" när man rör sig norrut genom Sverige.
Nu är förhållandena inte alltid så rätlinjiga och enkla som i gulärlans fall. Ofta har vi olika invandringsriktningar (och därmed flyttningsriktningar) för nordliga och sydliga populationer, tidtabeller som skiljer sig åt, olika genetik. Särskilt utpräglade kan sådana skillnader vara när man jämför sydsvenska populationer av arter, som i söder lägger två kullar, med mellansvenska populationer av samma arter, som inte hinner med mer än en kull. Det verkar finnas ett klimatiskt trappsteg nånstans i skarven Götaland-Svealand (eller rentav i norra Götaland), som återspeglas i helt skilda ruggningsförlopp för skånska och mellansvenska populationer av en och samma art. Vi fick korn på detta när vi i Fågelstudier (2:1-2, 3:2) jämförde ruggningsdiagram för sävsparv och gulärla från Skåne och Närke (vi skall arbeta upp dessa material och lägga in dem här nån gång när vi får tid). Här finns ett stort och mycket intressant undersökningsområde, där sydliga inlandsstationer som Landsjön, Tåkern och Hornborgasjön skulle kunna lämna viktiga bidrag! Kvismaren har ju redan gjort mycket, men säkert skulle det gå att få fram ännu mycket mer från det hållet.
HUR VI BEHANDLAR MATERIAL SOM VI FÅR IN FRÅN ANDRA RINGMÄRKARE
Om materialet är geografiskt isolerat, utan "grannar", bearbetas det isolerat, och presenteras i standardform med angivande av insamlingsperiod och den station/ringmärkare, som står för insamlandet. Där vi finner det lämpligt kommer vi att slå samman material - men naturligtvis i samförstånd med uppgiftslämnarna, och med angivande av alla som har lämnat bidrag.
NÅGRA REFERENSER:
Ginn, H.B. (1975) The timing and sequence of the complete annual moult in the Dunnock (Prunella modularis) in Britain over an eleven year period. Jour. Orn. 116: 263-280.
Ginn, H.B. & Melville, D.S. (1983) Moult in Birds. BTO GUIDE 19.
Hasselquist, D. et al. (1988) The seasonally divided feather moult in the Barred Warbler Sylvia nisoria - a new moult pattern for European passerines. Orn. Scand. 19: 280-286.
Haukioja, E. (1971) Processing moult card data with reference to the Chaffinch Fringilla coelebs. Orn. Fenn. 48: 25-32.
Holmberg, T. (1989) Blåhakens ruggning vid Ånnsjön. Fåglar i Jämtland-Härjedalen 9: 22-26.
Holmberg, T. (1992) The primary moult of the Brambling Fringilla montifringilla evaluated by four different methods. Orn. Sv. 2: 139-156.
Holmgren, N., Ellegren, H. & Pettersson, J. (1993) The adaptation of moult pattern in migratory Dunlins Calidris alpina. Orn.Scand. 24: 21-27.
Lundberg, P. & L.-O. Eriksson. (1984) Postjuvenile moult in two northern Scandinavian Starling Sturnus vulgaris populations - evidence for difference in the circannual time-program. Orn. Scand. 15: 105-109.
Ottosson, U. & F. Haas (1991) Primary moult of the Brambling Fringilla montifringilla in northern Sweden. Orn. Sv. 1: 113-118.
Sondell, J. (1977) Sävsparvens Emberiza schoeniclus ruggning i Kvismaren. Vår Fågelvärld 36: 174-184.
Sondell, J. (1987) Fyra trastarters ruggning i Kvismaren. Vår Fågelvärld 46: 55-63.
Sondell, J. Gulärlans ruggning i Kvismaren. Fågelstudier 3: 74-79.
Sondell, J. & Schildt, E.-B. (1985) Bofinkens ruggning i Kvismaren. Verks. vid Kvism. fågelst. 1984, s.2-8.
Ruggningsförloppet hos individ och population av törnsångare, lövsångare och gulärla i södra Skåne. Fågelstudier 2: 52-58.
Denna introduktion är skriven av Christer Persson, första version 15.5.98.