
|
|
Notes on biometry, sex determination and moult in Little Tern Sterna albifrons |
by Henning Behmann (Schleswig-Holstein/Germany) and Christer Persson (SW Scania/Sweden). With an appendix: Bird protection aspect of Little Tern ringing. |
| For readers from other continents: The Little Tern Sterna albifrons breeds throughout the southern Palearctic, north to Scotland and S Sweden, and has subspecific representatives in N America, Africa and Australia. There are three moult series in a season, but there is some ambiguity surrounding the extent of the second series. |
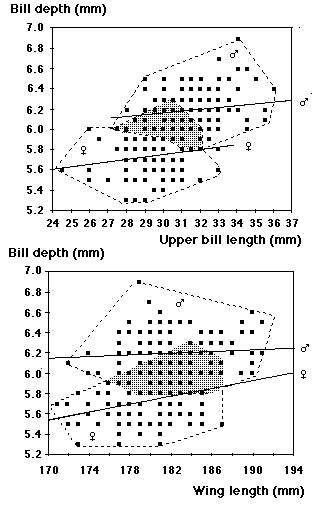
A majority of breeding Little Terns in N Europe can be sexed from the coloration of feet and tarsi in combination with the depth of the bill. The region of overlap in bill depth measured at the angle of the lower bill is 5.8 - 6.2 mm. In difficult cases the width of the loral line and the depth of the tail fork may be used as additional criteria. Breeding birds from the south Baltic had 2 - 5 new primaries from the third series in May - June. The number of black outer primaries was found to be age-independent, but slightly related to sex (more black primaries in females).
 Since 1965 one of us (HB) has ringed adult and young Little Terns in the Baltic coastal area of Schleswig-Holstein, Germany (Behmann, 1998). The main purpose has been to monitor breeding success and to protect the colonies of this endangered breeding species (category 2 on the Red List) of the Baltic area; as a by-product of the project an overwhelming amount of biometrical data has been collected. Recent measurements: 151 males from 1995 - 2001, 117 females from 1998 - 2001 constitute the material of this paper. In all years two Little Terns, one male, one female (out of 244 birds with known age handled between 1965 and 2001) were retrapped as breeding birds in their second calendar year, while 17 males, 13 females and 2 unsexed birds bred in their third calendar year, therefore unringed breeding birds caught on the nest have been aged 3c+ (although the majority probably does not breed until its fourth calendar year). In many birds the sex characters could be checked and substantiated during a sequence of years, and both contrahents of a pair were often caught together. Maximum wing length and tail fork were measured to the nearest millimetre, bill from feathers to the nearest half millimetre, bill depth at the angle of the lower bill to the nearest tenth of a millimetre. Unusual wear in remiges and broken bills were noted particularly, these measurements have been omitted in the calculations. Furthermore the moult status of primaries has been recorded. Our hope is, that this compilation will be of use to bird-ringers faced with the problems of sex-determination in colonies or at migration sites.
Since 1965 one of us (HB) has ringed adult and young Little Terns in the Baltic coastal area of Schleswig-Holstein, Germany (Behmann, 1998). The main purpose has been to monitor breeding success and to protect the colonies of this endangered breeding species (category 2 on the Red List) of the Baltic area; as a by-product of the project an overwhelming amount of biometrical data has been collected. Recent measurements: 151 males from 1995 - 2001, 117 females from 1998 - 2001 constitute the material of this paper. In all years two Little Terns, one male, one female (out of 244 birds with known age handled between 1965 and 2001) were retrapped as breeding birds in their second calendar year, while 17 males, 13 females and 2 unsexed birds bred in their third calendar year, therefore unringed breeding birds caught on the nest have been aged 3c+ (although the majority probably does not breed until its fourth calendar year). In many birds the sex characters could be checked and substantiated during a sequence of years, and both contrahents of a pair were often caught together. Maximum wing length and tail fork were measured to the nearest millimetre, bill from feathers to the nearest half millimetre, bill depth at the angle of the lower bill to the nearest tenth of a millimetre. Unusual wear in remiges and broken bills were noted particularly, these measurements have been omitted in the calculations. Furthermore the moult status of primaries has been recorded. Our hope is, that this compilation will be of use to bird-ringers faced with the problems of sex-determination in colonies or at migration sites.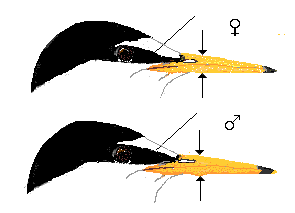

| Category | males: n | span (mm) | mean value ± 1 s.e.; 1 s.d. (mm) | females: n | span (mm) | mean value ± 1 s.e.; 1 s.d. (mm) |
|---|---|---|---|---|---|---|
| Wing-length, maximum chord | 151 | 172-191 | 182.6 ± 0.3; 3.9 | 117 | 171-187 | 179.9 ± 0.3; 3.5 |
| Upper bill from feathers | 151 | 27.5-36.0 | 31.6 ± 0.1; 1.6 | 117 | 24.5-33.0 | 29.3 ± 0.1; 1.4 |
| Lower bill from feathers | 151 | 28.5-36.5 | 31.7 ± 0.1; 1.5 | 116 | 25.5-32.0 | 29.5 ± 0.1; 1.3 |
| Black point, upper bill | 147 | 0-11.5 | 6.2 ± 0.2; 2.3 | 113 | 2-11 | 5.9 ± 0.2; 1.8 |
| Black point, lower bill | 146 | 0-11.5 | 5.0 ± 0.2; 2.1 | 112 | 0-9.0 | 4.6 ± 0.2; 1.8 |
| Bill depth | 151 | 5.8-6.9 | 6.20 ± 0.02; 0.19 | 117 | 5.3-6.3 | 5.73 ± 0.02; 0.20 |
| Tail fork | 134 | 29-53 | 40.7 ± 0.4; 4.4 | 84 | 24-48 | 34.6 ± 0.5; 4.9 |
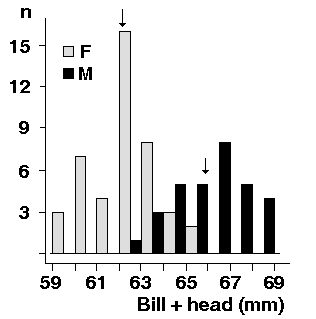
In contrast to the grey inner primaries, the two outermost ones are slate black in most cases. But field observations and museum skins show, that this is by no means always the case. There are more birds with one, three or four black primaries, and the number need not be the same in both wings. The number of black primaries probably declines with age. In juveniles all primaries are darker and the four or five outermost are deep black.What we feel, after reading and comparing the statements of different authors, is that this matter should be looked into again. Is it possible, that there are A./ black primaries - say, 7b or 8b - exchanged in series b? and B./ grey primaries - say, 7a or 8a - not exchanged in series b? We need to take a closer look at the primaries (using 5x or 10x magnification) in order to know if it is justified always to equal black primaries with primaries left unmoulted in series b. The field-season 2002 will be devoted to this task. So, what we describe in the following section is the number of black outer primaries, which may coincide with the number of primaries not moulted in series b, but on the other hand may fall below or even exceed this number. We have changed the text accordingly, writing "black" instead of "old" primaries.

Repeating: like the Roseate Tern Sterna dougallii and the White-winged Black Tern Chlidonias leucoptera, the Little Tern has three moult series; the last one (series c) running from February to April and comprising two to five inner primaries. When the birds are brooding or feeding young or when young are fledged, a new series (series a), starting with the innermost primary (P1), is initiated:
1. 3c+ brooding female, Fastensee, Fehmarn 17.6.01. P1 vane emerging from sheath, P2 pin, P3 - 5 fresh from series 3 - i.e. maximum performance in moult series 3 as well!In May and June 2001 - 02 we recorded the moult status of adult breeding birds in both Schleswig-Holstein and in SW Scania, the number of fresh primaries from the 3rd series is given in Table II:
2. 16c+ brooding female, Bottsand, Bay of Kiel, Germany 15.6.02. P1 0.5.
3. 14c+ brooding male, 19.6.02, Germany. P1 shed
4. 11c brooding male, 22.6.02, Germany. P1 shed
5. 5c+ brooding female, 23.6.02, Germany. P1 - 2 shed in one wing, P1 shed in the other.
6. 14c+ brooding male, 29.6.02, Germany. P1 shed
| Germany; new primaries | n, 2001 | n, 2002 | Scania; new primaries | n, 2001 | n, 2002 |
|---|---|---|---|---|---|
| P1 - P2 | 5 | 4 | P1 - P2 | - | - |
| P1 - P3 | 43 | 21 | P1 - P3 | 12 | 8 |
| P1 - P4 | 25 | 20 | P1 - P4 | 2 | 7 |
| P1 - P5 | 2 | 5 | P1 - P5 | - | - |
| Germany + Scania new primaries | n, 2001 | n, 2002 |
|---|---|---|
| P1 - P2 or P1 - P3 | 60 | 33 |
| P1 - P4 or P1 - P5 | 29 | 32 |
7. 3c+, accompanied by young, Skanör 14.8.99. P1, P2 fresh, P3 0.5, rest of primaries older; ten old tail-feathers.In subsequent years three birds conducting juveniles could be sexed, all were females. The moult state of no. 9 suggests suspension after P6 - before the large primaries 7 - 10 (which are then the first to be moulted on wintering grounds) - and consecutive migration, other late birds must suspend earlier or migrate with growing wing:
8. 3c+, accompanied by young, Skanör 17.8.99. P1 - 3 "fresh", P4 - 6 worn, P7 - 10 very worn. (A writing error corrected 7.9.01)
9. 3c+ female, accompanied by two young, Skanör 15.8.01. P1 - 4 fresh, P5 0.9, P6 short pin; P9-10 black (from 2nd series?). S1 fresh, S2 0.4; R1 fresh, R2 0.6.
10. 3c+ female, accompanied by two young, Skanör 17.8.01. P1 fresh, P2 0.7, rest of primaries older; P9-10 black (from 2nd series?), ten old tail-feathers.
11. 3c+ female, accompanied by at least one young, Skanör 18.8.03. P1 - 2 fresh, P3 0.8; P7 - 8 less worn than P4, 5, 6, 9 and 10.
The second moult series is interrupted when breeding starts in May, in most cases the process leaves at least the two outermost primaries (P9 and P10) unmoulted. These primaries have a blackish and worn look, contrasting with silvery, rather fresh primaries 7 and 8 (but we need to investigate if there are silvery, unmoulted primaries). A bird in its 18th calendar year from the Falsterbo peninsula 25.5.01 had all outer primaries silvery and fresh (the only one with distinct silvery hue on outer primaries so far seen in Scania), while a bird in its 11th calendar year had P8 - 10 black, suggesting that the number of black primaries could be age-dependent - as suggested by Nadler (1995). The material from Schleswig-Holstein 1995 - 2001 comprises 170 birds; 83 ringed as breeding birds (in their 3rd calendar year or older), 87 ringed as pulli (i.e. of known age when retrapped in later years). Analyzing the "moult cards" of birds of known age (3c - 16c) we tested for age-dependence in the number of black primaries, numbers are given in Table IV:| Age category | P10; n | P10/P10-P9; n | P10-P9; n | P10-P9/P10-P8; n | P10-P8; n |
|---|---|---|---|---|---|
| 3c - 6c | 5 | 1 | 19 | 3 | 3 |
| 7c - 10c | 6 | 0 | 22 | 0 | 2 |
| 11c - 16c | 7 | 1 | 13 | 1 | 4 |
Next we test for the influence of sex on the number of black primaries. Here the whole material (103 males, 67 females) can be used, numbers are presented in Table V:| Sex | P10 or P10/P10-P9; n | P10-P9 or P10-P9/P10-P8; n | P10-P8; n |
|---|---|---|---|
| Male | 23 | 70 | 10 |
| Female | 7 | 46 | 14 |
In Scania, many breeding sites are more "natural" than in Germany, and the whole production often gets predated by Herring Gulls and Great Black-backed Gulls. CP has tried the following protection measures: