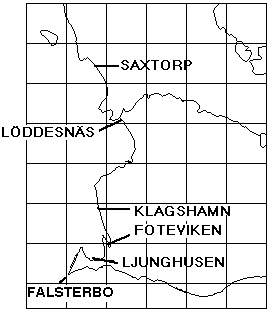
Fig. 1. Location of major roosts, where Yellow Wagtails have been ringed 1964-99. Many birds leave Sweden by way of "Nabben", the SW cape of the Falsterbo peninsula.
Introduction: A long-term study of Yellow Wagtails (Motacilla flava) at roosts in SW Scania, Sweden |
| For readers from other continents: The Yellow Wagtail Motacilla flava breeds with numerous subspecies throughout the Palearctic. The subspecies flava breeds in the southern half of Sweden, in southern Finland and in southernmost Norway, the subspecies thunbergi in northern Sweden, northern Finland and most of Norway, in addition there are hybrids in a zone of encounter in all three countries. Adults moult completely in late summer and the end of moult is exactly synchronized with the start of migration. |
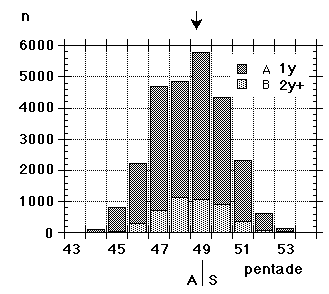
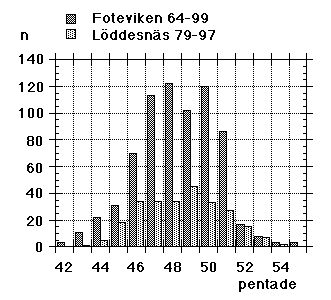
 The synchronism and coordination on the autumn side of the annual cycle have been main topics of interest in a long-term study (1964-99), where large numbers of migrating Yellow Wagtails were caught, ringed and investigated for moult at roosts in SW Scania. In addition the same work has provided information on breeding and wintering areas as well as on migration routes of the populations involved; other conditions underlying the general stability can be assessed against this background. So, this paper deals with interaction, between physiological processes, between populations, between different segments of the annual cycle, and the overall issue is the relative stability resulting from this interaction (and synchronous action) at different levels.Between the Foteviken area and Klagshamn (c55� 31', 12� 56'), there are about 25 separate reeds that will serve as roosts for Yellow Wagtails. (Fig. 1) The actual choice - with new point of gravity from night to night - is decided mainly by wind-direction, the descending flocks always approaching and entering the reed-beds against the wind. The presence of predators will affect the choice to some extent as well. By the end of August as much as 10.000 Yellow Wagtails will spend the night in the overall area. Given the shifting preferences, a tape lure is very helpful if the ringer wants to catch as many birds as possible, in addition the birds will be caught earlier in this way, and daylight always facilitates both handling and race determination. Still, a tape lure was not consistently used at Foteviken until 1986, so the material collected here before that year is "undisturbed" by this remedy. In contrast, tape lures were used at Löddesnäs from the very first year, and this is reflected in catching numbers; they are close to the optimum of the ringer, reduced only by periods of bad weather. The setback with lures is that catching numbers will not reflect the number of birds spending the night in an area; if the nets are properly set the catch will at best reflect the number of nets used. With a lure it is as easy to catch 300 birds out of a thousand as it is to catch them out of ten thousand (or even easier).In the Foteviken area 53.100 Yellow Wagtails were caught between 1964 and 1999; 1.582 not properly aged between 1964 and 1966, from 1967 onwards 1.060 Motacilla flava flava in nest plumage, 41.744 "plain" juveniles, mostly in 1st winter plumage, 7.426 adults and 1.288 unaged birds. Christer Persson (with many helpers) started this catch in 1964, Peter Olsson and Kerstin Norrman joined from 1986-87 and made independent contributions in the early nineties, they in turn assisted by Per Nothagen; in the late nineties the catching effort gradually faded away. By then Yellow Wagtails had been caught on 714 separate occasions, some hundred of these including only a couple of birds.Parallel to this, Ulf Lundwall started ringing Yellow Wagtails at the mouth of River Lödde (Löddesnäs, see Fig. 1) in 1979; up till and including 1997 (aided by Håkan Skoglund, in addition Peter Olsson ringed here on five occasions in 1995) this resulted in 25.978 birds ringed on 255 occasions: 21.241 juveniles, 4.731 adults and 6 unaged. Unlike Foteviken/Klagshamn Löddesnäs is one restricted reed, serving as roost for many species, including Starlings (Sturnus vulgaris) and hirundines, from this point of view it is the better site; single catches of 300 birds are easily obtained here with little more than ten nets. It should be added, that in many years activity at Löddesnäs was restricted to weekends (Friday - Sunday), while Foteviken ringers have had more freedom to select the days or periods of peak migration.All in all 79.078 Yellow Wagtails were ringed in 36 years at these two sites (i.e. an average of more than 2.000 per year over a period of three and a half decades), resulting in 173 long-distance recoveries (0.22 %) and some 500 local retraps within and between roosts. In addition 9 birds with foreign rings were controlled, and 17 birds ringed elsewhere in Sweden. Furthermore 506 moult-cards related to actual postnuptial moult (from first remige shed to sheath on ninth primary and inner secondaries) were collected at Foteviken, Klagshamn and Löddesnäs. These combined data: distributions of race- and age-determined birds, recoveries and moult cards form the material of this paper.
The synchronism and coordination on the autumn side of the annual cycle have been main topics of interest in a long-term study (1964-99), where large numbers of migrating Yellow Wagtails were caught, ringed and investigated for moult at roosts in SW Scania. In addition the same work has provided information on breeding and wintering areas as well as on migration routes of the populations involved; other conditions underlying the general stability can be assessed against this background. So, this paper deals with interaction, between physiological processes, between populations, between different segments of the annual cycle, and the overall issue is the relative stability resulting from this interaction (and synchronous action) at different levels.Between the Foteviken area and Klagshamn (c55� 31', 12� 56'), there are about 25 separate reeds that will serve as roosts for Yellow Wagtails. (Fig. 1) The actual choice - with new point of gravity from night to night - is decided mainly by wind-direction, the descending flocks always approaching and entering the reed-beds against the wind. The presence of predators will affect the choice to some extent as well. By the end of August as much as 10.000 Yellow Wagtails will spend the night in the overall area. Given the shifting preferences, a tape lure is very helpful if the ringer wants to catch as many birds as possible, in addition the birds will be caught earlier in this way, and daylight always facilitates both handling and race determination. Still, a tape lure was not consistently used at Foteviken until 1986, so the material collected here before that year is "undisturbed" by this remedy. In contrast, tape lures were used at Löddesnäs from the very first year, and this is reflected in catching numbers; they are close to the optimum of the ringer, reduced only by periods of bad weather. The setback with lures is that catching numbers will not reflect the number of birds spending the night in an area; if the nets are properly set the catch will at best reflect the number of nets used. With a lure it is as easy to catch 300 birds out of a thousand as it is to catch them out of ten thousand (or even easier).In the Foteviken area 53.100 Yellow Wagtails were caught between 1964 and 1999; 1.582 not properly aged between 1964 and 1966, from 1967 onwards 1.060 Motacilla flava flava in nest plumage, 41.744 "plain" juveniles, mostly in 1st winter plumage, 7.426 adults and 1.288 unaged birds. Christer Persson (with many helpers) started this catch in 1964, Peter Olsson and Kerstin Norrman joined from 1986-87 and made independent contributions in the early nineties, they in turn assisted by Per Nothagen; in the late nineties the catching effort gradually faded away. By then Yellow Wagtails had been caught on 714 separate occasions, some hundred of these including only a couple of birds.Parallel to this, Ulf Lundwall started ringing Yellow Wagtails at the mouth of River Lödde (Löddesnäs, see Fig. 1) in 1979; up till and including 1997 (aided by Håkan Skoglund, in addition Peter Olsson ringed here on five occasions in 1995) this resulted in 25.978 birds ringed on 255 occasions: 21.241 juveniles, 4.731 adults and 6 unaged. Unlike Foteviken/Klagshamn Löddesnäs is one restricted reed, serving as roost for many species, including Starlings (Sturnus vulgaris) and hirundines, from this point of view it is the better site; single catches of 300 birds are easily obtained here with little more than ten nets. It should be added, that in many years activity at Löddesnäs was restricted to weekends (Friday - Sunday), while Foteviken ringers have had more freedom to select the days or periods of peak migration.All in all 79.078 Yellow Wagtails were ringed in 36 years at these two sites (i.e. an average of more than 2.000 per year over a period of three and a half decades), resulting in 173 long-distance recoveries (0.22 %) and some 500 local retraps within and between roosts. In addition 9 birds with foreign rings were controlled, and 17 birds ringed elsewhere in Sweden. Furthermore 506 moult-cards related to actual postnuptial moult (from first remige shed to sheath on ninth primary and inner secondaries) were collected at Foteviken, Klagshamn and Löddesnäs. These combined data: distributions of race- and age-determined birds, recoveries and moult cards form the material of this paper.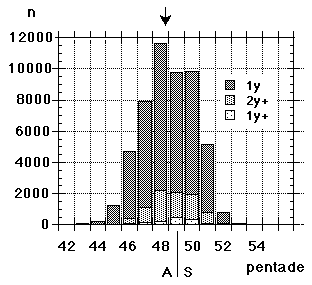
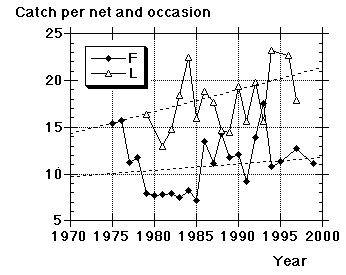
| Year, site | Total | 1y | 2y+ | 1y+ | Main interval | No. of catches in main interval | Average catch/occasion in main interval | Average no. of nets (no. of occasions) |
|---|---|---|---|---|---|---|---|---|
| FOV, 1967 | 1035 | 895 | 140 | 69 | 22.8-12.9 | 13 | 75 | 7.0 (7) |
| FOV, 1968 | 904 | 388 | 37 | 479 | 20.8-6.9 | 10 | 82 | - |
| FOV, 1969 | 1138 | 803 | 130 | 205 | 19.8-12.9 | 12 | 87 | - |
| FOV, 1970 | 1390 | 1241 | 144 | 5 | 17.8-11.9 | 18 | 72 | - |
| FOV, 1971 | 843 | 722 | 118 | 3 | 19.8-13.9 | 17 | 46 | - |
| FOV, 1972 | 665 | 578 | 52 | 35 | 28.8-11.9 | 12 | 53 | - |
| FOV, 1973 | 1723 | 1530 | 192 | 1 | 15.8-8.9 | 23 | 74 | - |
| FOV, 1974 | 1314 | 1063 | 229 | 22 | 19.8-10.9 | 17 | 76 | - |
| FOV, 1975 | 2200 | 1881 | 305 | 20 | 14.8-9.9 | 20 | 104 | 6.8 |
| FOV, 1976 | 2437 | 2242 | 176 | 19 | 14.8-12.9 | 24 | 97 | 6.2 |
| FOV, 1977 | 1913 | 1548 | 336 | 29 | 15.8-11.9 | 23 | 82 | 7.3 |
| FOV, 1978 | 925 | 823 | 94 | 8 | 16.8-8.9 | 10 | 90 | 7.6 |
| LNÄ1 1979 FOV 1979 | 428 | 369 | 59 | - | 25.8-4.9 | 4 | 107 | 6.5 |
| 902 | 789 | 110 | 3 | 17.8-9.9 | 15 | 58 | 7.3 | |
| LNÄ 1980 FOV 1980 | 482 | 452 | 30 | - | 1.9-10.9 | 5 | 97 | 7.4 |
| 927 | 800 | 117 | 4 | 25.8-11.9 | 16 | 58 | 7.5 | |
| LNÄ 1981 FOV 1981 | 1157 | 907 | 250 | - | 7.8-16.9 | 11 | 104 | (11.3)2 |
| 1066 | 830 | 222 | 14 | 19.8-9.9 | 15 | 69 | 8.9 | |
| LNÄ 1982 FOV 1982 | 937 | 820 | 117 | - | 15.8-5.9 | 7 | 133 | 9.0 |
| 1580 | 1154 | 394 | 32 | 16.8-10.9 | 18 | 86 | 10.8 | |
| LNÄ 1983 FOV 1983 | 2033 | 1737 | 296 | - | 17.8-14.9 | 13 | 152 | 8.3 |
| 1892 | 1575 | 287 | 30 | 15.8-14.9 | 22 | 84 | 11.2 | |
| LNÄ 1984 FOV 1984 | 1625 | 1286 | 338 | 1 | 9.8-10.9 | 13 | 124 | 5.5 |
| 1471 | 1062 | 341 | 68 | 20.8-10.9 | 14 | 104 | 12.6 | |
| LNÄ 1985 FOV 1985 | 1861 | 1458 | 403 | - | 15.8-2.9 | 15 | 97 | 6.1 |
| 1099 | 885 | 208 | 6 | 19.8-10.9 | 17 | 64 | 8.9 | |
| LNÄ 1986 FOV 1986 | 2281 | 1994 | 287 | - | 11.8-13.9 | 18 | 126 | 6.7 |
| 3775 | 3110 | 630 | 35 | 13.8-13.9 | 30 | 126 | 9.3 | |
| LNÄ 1987 FOV 1987 | 2750 | 2098 | 648 | 4 | 16.8-8.9 | 17 | 161 | 9.1 |
| 2658 | 2094 | 546 | 18 | 16.8-13.9 | 23 | 114 | 10.3 | |
| LNÄ 1988 FOV 1988 | 2837 | 2321 | 516 | - | 14.8-17.9 | 21 | 135 | 9.2 |
| 4914 | 4147 | 750 | 17 | 9.8-17.9 | 33 | 147 | 10.3 | |
| LNÄ 1989 FOV 1989 | 1417 | 1159 | 258 | - | 13.8-14.9 | 13 | 108 | 7.5 |
| 4504 | 3851 | 615 | 38 | 10.8-14.9 | 31 | 141 | 12.0 | |
| LNÄ 1990 FOV/KLH 1990 LJH 1990 | 1063 | 911 | 152 | - | 13.8-5.9 | 8 | 132 | 6.8 |
| 2641 | 2417 | 213 | 11 | 10.8-10.9 | 27 | 97 | -3 | |
| 898 | 823 | 64 | 11 | 17.8-12.9 | 20 | 45 | 4.9 | |
| LNÄ 1991 FOV/KLH 1991 | 1030 | 848 | 182 | - | 20.8-9.9 | 8 | 123 | 8.0 |
| 880 | 764 | 108 | 8 | 26.8-13.9 | 12 | 73 | 7.9 | |
| LNÄ 1992 FOV 1992 | 872 | 669 | 203 | - | 18.8-10.9 | 8 | 107 | 5.4 |
| 1140 | 957 | 174 | 9 | 14.8-9.9 | 10 | 111 | 8.0 | |
| LNÄ 1993 FOV/KLH 1993 | 637 | 544 | 93 | - | 22.8-12.9 | 7 | 89 | 5.7 |
| 1455 | 1158 | 219 | 78 | 15.8-16.9 | 13 | 107 | 6.1 | |
| LNÄ 1994 FOV 1994 | 1642 | 1332 | 310 | - | 17.8-7.9 | 11 | 146 | 6.3 |
| 1277 | 1054 | 211 | 12 | 10.8-17.9 | 15 | 82 | 7.6 | |
| LNÄ 1995 FOV 1995 | 497 | 368 | 128 | 1 | 17.8-19.9 | 6 | 83 | - |
| 738 | 651 | 81 | 6 | 16.8-12.9 | 9 | 81 | 7.1 | |
| LNÄ 1996 | 971 | 797 | 174 | - | 25.8-14.9 | 7 | 136 | 6.0 |
| LNÄ 1997 FOV 1997 | 1458 | 1171 | 287 | - | 13.8-3.9 | 11 | 131 | 7.3 |
| 593 | 521 | 67 | 5 | 21.8-31.8 | 7 | 85 | 6.7 | |
| FOV 1999 | 427 | 376 | 49 | 2 | 20.8-10.9 | 6 | 69 | 6.2 |
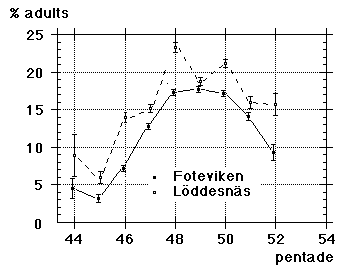
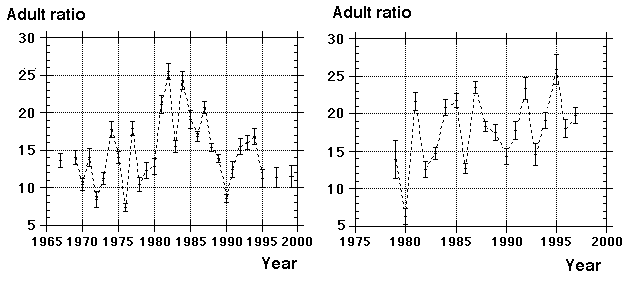 |
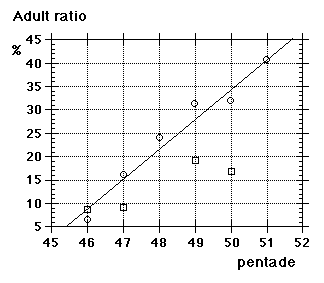
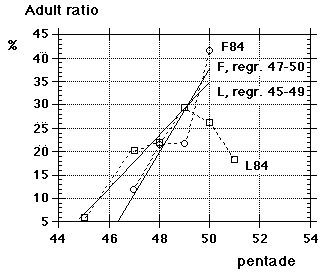
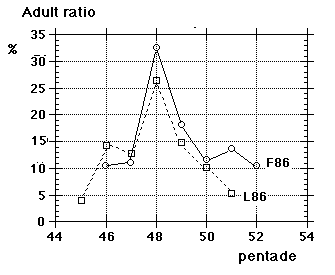
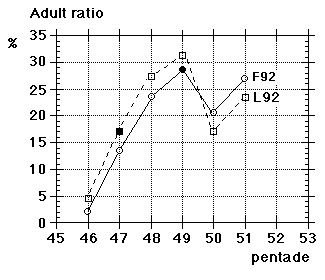
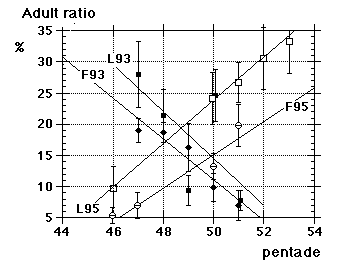
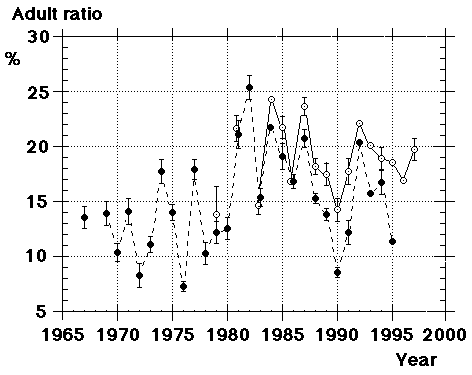
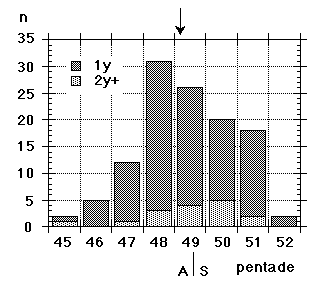
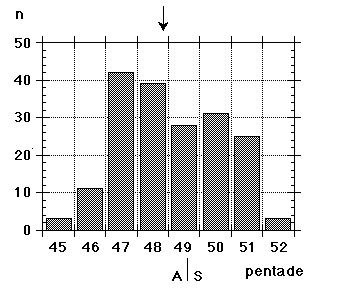
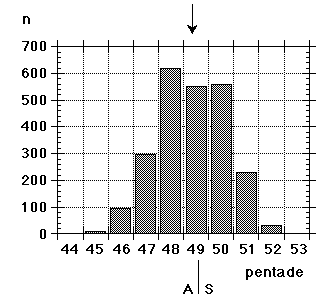
 Unfortunately we did not note "southern" flava separately from the beginning, although we could recognize them; all "flava type" birds went under one single heading between 1967 and 1986. Instead the first step was to isolate the obvious hybrid flava/thunbergi; it was noted separately at Foteviken from 1967, and our own classification could take support and guidance from Sammalisto (1968). After that we gradually began to separate "southern" and "northern" flava, and these distinctions were put into practice in 1987 and 1988. In the latter cases we concentrated on adult males, females do not "suggest" their origin in the way males do. Finally, from 1988 onwards, we were confronted with green-headed and green-backed juveniles, strongly reminding of the subspecies flavissima, many of them even with yellow tone in their supercilium (1y birds of this subspecies are said to have buffish-white or white supercilium). Any remaining doubt was cleared away by the first adult male; up till and including 1994 there followed fifteen more adults, in addition 100 "possible" juvenile flavissima were ringed. In addition two adult males of feldegg type were caught, one at Foteviken 10.9.91, one at Löddesnäs 27.8.93. So, in our heyday, we discerned seven different main forms in South Swedish roosts, the type appearance of males in winter plumage is shown in Fig. 15:
Unfortunately we did not note "southern" flava separately from the beginning, although we could recognize them; all "flava type" birds went under one single heading between 1967 and 1986. Instead the first step was to isolate the obvious hybrid flava/thunbergi; it was noted separately at Foteviken from 1967, and our own classification could take support and guidance from Sammalisto (1968). After that we gradually began to separate "southern" and "northern" flava, and these distinctions were put into practice in 1987 and 1988. In the latter cases we concentrated on adult males, females do not "suggest" their origin in the way males do. Finally, from 1988 onwards, we were confronted with green-headed and green-backed juveniles, strongly reminding of the subspecies flavissima, many of them even with yellow tone in their supercilium (1y birds of this subspecies are said to have buffish-white or white supercilium). Any remaining doubt was cleared away by the first adult male; up till and including 1994 there followed fifteen more adults, in addition 100 "possible" juvenile flavissima were ringed. In addition two adult males of feldegg type were caught, one at Foteviken 10.9.91, one at Löddesnäs 27.8.93. So, in our heyday, we discerned seven different main forms in South Swedish roosts, the type appearance of males in winter plumage is shown in Fig. 15: |
 |
 |
 |
 |
 |
 |
 |
|
 |
|
 |
|
 |
|
 |
|
 |
|
 |
|
 |
|